






200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过





骨质疏松症是一种以骨量低下、骨组织微结构损坏,导致骨脆性增加,易发生骨折为特征的全身性骨病[1],已成为我国重要的公共卫生问题之一。药物治疗是抗骨质疏松症的重要手段。今年发布的《原发性骨质疏松症诊疗指南(2022版)》[1]新增推荐了艾地骨化醇等药物,丰富了选择,优化了格局。作为新型活性维生素D类似物,艾地骨化醇较传统活性维生素D在分子结构、作用机制、临床疗效方面都有着独到优势。
本期“骨舞健言”有幸邀请到山东大学骨质疏松与骨矿盐疾病研究中心主任、山东大学口腔医院骨代谢研究室主任李敏启教授接受专访,从基础和临床角度全方位介绍了艾地骨化醇的特点,并展望了其广阔的探索前景,帮助大家深刻认识这一创新药物的价值。本文整理访谈精要如下。
李敏启 教授
口腔医学博士、山东大学教授、博士生导师
山东省泰山学者特聘专家
山东大学齐鲁青年学者
日本国立北海道大学特聘教授
山东大学骨质疏松与骨矿盐疾病研究中心主任
山东大学口腔医学院骨代谢研究室主任
深耕艾地骨化醇相关基础研究多年
点击即可观看
您长期从事艾地骨化醇相关的基础研究工作并取得了一系列重大成果,这最初源于什么样的契机?同时请您介绍一下艾地骨化醇的分子结构特点和研发历程。
李敏启教授:我与艾地骨化醇的“缘分”可以追溯到2006年,至今已经17载。上个世纪末我赴日本留学后,在日本工作了很长一段时间,期间有幸参与到了艾地骨化醇的研发过程中,是机缘巧合,也是专业使然。
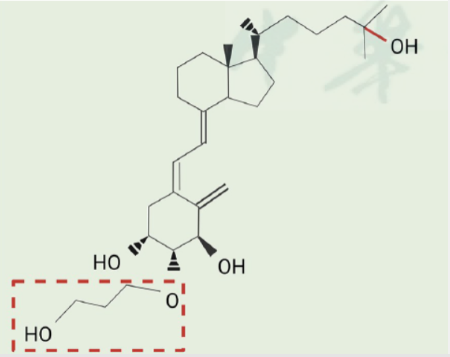
艾地骨化醇源于活性维生素D,其在骨化三醇基础上创新性地于2β位添加了羟基丙氧基,成为一种新型的活性维生素D类似物[1α,25-(OH)2-2β-(3-羟基丙氧基)维生素D3,图1][2]。3-羟基丙氧基这一小基团赋予了艾地骨化醇多维度的优势。
首先,艾地骨化醇与血清中的维生素D结合蛋白(vitamin D binding protein, DBP)结合能力较强,实现了长半衰期。
其次,尽管艾地骨化醇与维生素D受体(vitamin D receptor, VDR)的亲和力不是很高,但其3-羟基丙氧基可通过H键和CH-π相互作用使艾地骨化醇-VDR二元复合物非常稳定,改善了结合动力学,使之具有良好的药理活性[3]。
再者,由于肾脏24-羟化酶(CYP24A1)中没有3-羟基丙氧基的空间,因此艾地骨化醇不会被肾脏的CYP24A1快速降解,其稳定性从而增强;且艾地骨化醇的代谢也由传统活性维生素D的肾脏依赖性转变为肝脏依赖性,大大减少了骨质疏松患者的肾脏代谢负担[3]。
得益于分子结构的创新,艾地骨化醇较传统活性维生素D拥有了这些独特的药学和机制优势,为优异的临床疗效奠定了基石。
图1 艾地骨化醇分子式[2]
回顾艾地骨化醇的研发历程,从首次发现至今已近四十年,走过了坚实的循证之路。
1985年,艾地骨化醇被发现,当时命名为“ED-71”。
2004年,艾地骨化醇在日本的Ⅲ期临床试验启动。研究证实,相比阿法骨化醇,艾地骨化醇使新椎体骨折发生风险降低了26%;且艾地骨化醇在早期即具有腕部骨折抑制效果,36个月的骨折发生风险降低达71%。该研究结果于2011年发表在国际知名期刊Bone杂志[4]。
2011年,即研究结果发表同年,艾地骨化醇获得日本厚生劳动省批准用于防治骨质疏松症。
2015年,艾地骨化醇在中国的Ⅲ期临床试验启动,研究设计与日本的研究相似。结果显示,与阿法骨化醇组相比,艾地骨化醇组的腰椎、全髋和股骨颈骨密度分别增加了2.05%、1.33%和1.78%。研究全文于2019年发表在Journal of Bone and Mineral Metabolism [5]。
2020年,艾地骨化醇获得了国家药品监督管理局颁发的进口药品注册证,被批准用于我国绝经后女性骨质疏松症患者治疗。
可以看到,艾地骨化醇的机制优势在临床试验中得到了充分体现,较传统的活性维生素D药物显著提升了骨质疏松症防治效果。
您所在的课题组于2011年首次发现了艾地骨化醇诱导“Mini-modeling”骨形成现象,这一骨形成过程与“传统骨改建骨形成”有何不同?艾地骨化醇在诱导“Mini-modeling”骨形成改善骨质疏松方面的作用机制有哪些?
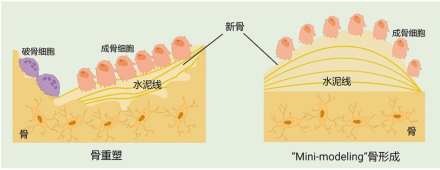
我们课题组在2011年首次发现了艾地骨化醇诱导的“Mini-modeling”骨形成现象,即在艾地骨化醇治疗的卵巢切除(OVX)大鼠骨小梁表面观察到了大量以小山丘样的“骨芽”形态存在的“Mini-modeling”骨形成(图2),骨基质表面聚集了大量形态丰满的成熟成骨细胞[7]。
“Mini-modeling”骨形成的机制与“传统骨改建骨形成”有着本质的区别,简言之,后者是先“破坏”再“重建”,而前者无需“破坏”、直接“新建”,直接“新建”的骨质在形态和“质地”方面都有着明显优势。具体而言,“Mini-modeling”骨形成不需要破骨细胞骨吸收这个环节,而是直接通过一些因素刺激覆盖在骨基质表面的静止的、扁平的前成骨细胞增殖分化为成熟的成骨细胞,将新产生的骨基质直接沉积在既存骨组织表面形成新骨。因此,传统骨改建骨形成过程是先形成一个骨基质陷窝,然后在其中填充新骨;而“Mini-modeling”骨形成是在原来的骨基质表面连续形成形态微小、水泥线较为平滑的新骨[8-9]。
图2 艾地骨化醇诱导不依赖于破骨细胞骨吸收的“Mini-modeling”骨形成[2]
基于艾地骨化醇诱导“Mini-modeling”骨形成这一现象的发现,我们课题组对其改善骨质疏松的具体作用继续展开了深入探索,目前主要揭示了以下几方面机制。
其一,艾地骨化醇能够通过增强碱性磷酸酶的活性以及活化P13K/AKT信号通路,促进骨髓间充质干细胞(BMSCs)的成骨向分化[10]。同时我们也发现,给予艾地骨化醇治疗后,OVX大鼠因雌激素丢失所致的BMSCs衰老得到了逆转和抑制、成骨分化能力增强,而这一作用可能是通过调控氧化应激来实现的。这就从BMSCs衰老角度解释了艾地骨化醇治疗骨质疏松症的部分作用机制[11]。
其二,既往研究表明,骨细胞表达VDR,而活性维生素D能够通过调控骨细胞功能影响骨改建[12-13]。本课题组前期研究发现艾地骨化醇诱导的“Mini-modeling”新生骨基质中骨细胞分泌的硬骨素明显减少,而硬骨素与骨形成之间的关系非常密切。所以我们认为骨细胞可能在艾地骨化醇诱导的“Mini-modeling”骨形成过程中发挥了重要调控作用,相关机理仍在进一步探究。
其三,从免疫角度,我们近期明确了艾地骨化醇能够调控适应性免疫中CD4+辅助T细胞亚群的平衡,进而创造更有利于骨形成的微环境。因此,骨免疫学相关研究也可能为阐明艾地骨化醇诱导“Mini-modeling”骨形成提供新的思路。
相信艾地骨化醇诱导“Mini-modeling”骨形成改善骨质疏松的机制远不止于上述方面,其作用是多维度的,需要更多的基础和转化研究去发掘。
《原发性骨质疏松症诊疗指南(2022版)》[1]中指出,艾地骨化醇治疗期间,在常规饮食情况下可不必服用钙剂。请您从机制的角度解读这一推荐背后的原因。
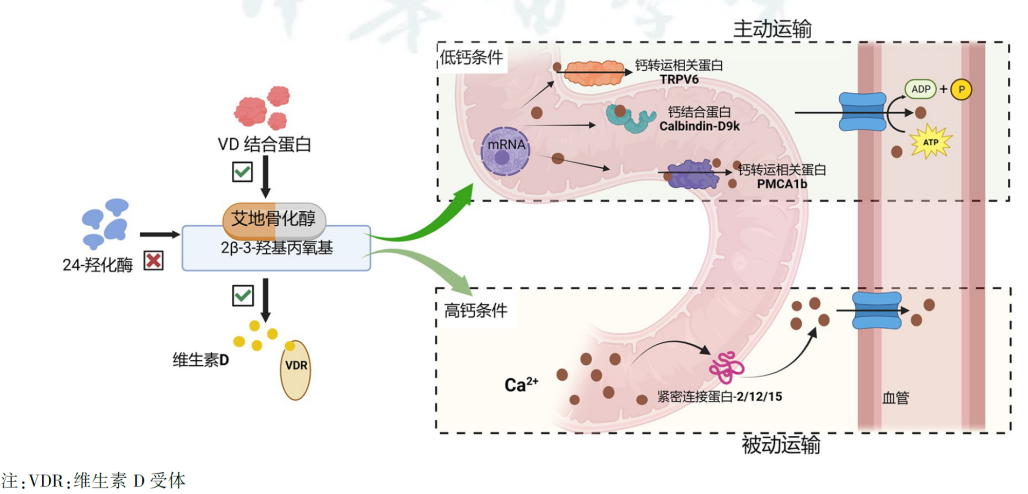
传统活性维生素D主要通过两方面机制促进钙吸收,一方面促进钙转运相关蛋白(TRPV6、钙结合蛋白、PMCA1b等)的生物合成,并增加钙的通透性促进钙离子主动向血液转运;同时可上调紧密连接蛋白-2/12/15的表达,促进钙的被动运输[14]。
相比传统活性维生素D,艾地骨化醇可从多角度增强肠道吸收钙的能力(图3)。
首先,艾地骨化醇的3-羟基丙氧基与DBP的结合力较强,具有较长的血浆半衰期。
其次,3-羟基丙氧基阻碍了艾地骨化醇与CYP24A1的结合,使其不易被代谢失活,可更持久地在靶细胞内发挥活性[15-16]。
此外,艾地骨化醇能够增加TRPV6及钙结合蛋白Calbindin-D9K在十二指肠的mRNA表达,更有效地促进钙的主动运输,提高肠道钙吸收率[17]。
图3 艾地骨化醇通过2β-3-羟基丙氧基促进肠道钙吸收[2]
我们在Ⅲ期临床试验中也观察到,艾地骨化醇在没有额外补充钙剂的情况下,很好地改善了骨质疏松,从临床角度充分印证了其能够有效增强肠道钙吸收的作用,也为指南推荐提供了依据。
请您对艾地骨化醇抗骨质疏松症的作用机制进行简要总结,并谈谈艾地骨化醇其他作用机制的研究方向。
李敏启教授:艾地骨化醇作为一种新型活性维生素D类似物,在改善骨质疏松、预防骨折并发症方面疗效显著持久,主要作用机制(图4)包括:
第一,通过鞘氨醇1磷酸(sphingosine-1-phosphate, S1P)机制及EphrinB2/EphB4信号通路调控破骨细胞分化而有效抑制骨吸收;
第二,通过诱导独特的“Mini-modeling”骨形成机制,有效促进骨形成和骨基质矿化,增加骨小梁的连接性、提升骨密度和改善骨结构;
第三,通过有效促进钙的主动运输,显著提高肠道钙吸收率。
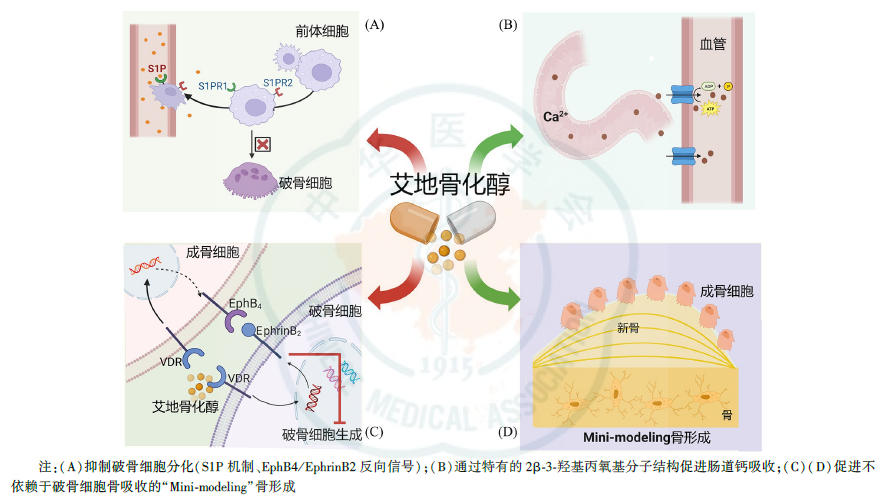
另外,我们课题组也在持续探索艾地骨化醇在其他领域疾病的作用,希望进一步拓展其应用范围、使更多患者受益。例如神经系统领域的阿尔茨海默症、精神分裂症,自身免疫相关的糖皮质激素性骨质疏松症(GIOP),口腔领域的牙周炎、糖尿病性牙周炎,以及骨质疏松与糖尿病的相互影响等。期待艾地骨化醇在更多疾病领域取得突破,不断释放潜力,发挥更大临床价值!
参考文献:
[1]中华医学会骨质疏松和骨矿盐疾病分会.中华骨质疏松和骨矿盐疾病杂志,2022,15(6):573-611.
[2]李敏启. 中华内分泌代谢杂志, 2023, 39(8) : 726-732.
[3]Ono Y. J Steroid Biochem Mol Biol,2014,139:88-97.
[4]Matsumoto T,et al. Bone,2011,49(4):605-612.
[5]Jiang Y, et al. J Bone Miner Metab,2019,37(6):1036-1047.
[6]Matsuo K, Irie N. Arch Biochem Biophys,2008,473(2):201-209.
[7]de Freitas PH, et al. Bone,2011,49(3):335-342.
[8]Jee WS, et al. J Musculoskelet Neuronal Interact,2007,7(3):232-239.
[9]Yamamoto T, et al. Biomed Res,2021,42(5):161-171.
[10]Lu Y, et al. Stem Cell Res Ther,2022,13(1):113.
[11]Kou Y, et al. Front Pharmacol,2023,14:1067085.
[12]Rolvien T, et al. Bone,2017,103:78-87.
[13]Wang Y, et al. J Bone Miner Res,2014,29(3):685-692.
[14]Christakos S, et al. Mol Cell Endocrinol,2011,347(1-2):25-29.
[15]Kondo S, et al. J Steroid Biochem Mol Biol,2015,148:232-238.
[16]Ritter CS, Brown AJ. J Cell Biochem,2011,112(5):1348-1352.
[17]Hirota Y, et al. PLoS One,2018,13(10):e0199856.
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017