









200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过




陈晶蕾 综述 鲍 浩 刘志红 审校
DOI:10.3969/j.issn.1006-298X.2025.06.013
[基金项目]江苏省卫生健康委员会医学科研重点项目(K2023003);南京大学医学院教学改革课题(NDY2025004)
[作者单位]南京大学医学院附属金陵医院(东部战区总医院) 博士研究生(陈晶蕾) 国家肾脏疾病临床医学研究中心(南京,210016)
摘 要 CD36作为一种多功能受体,可识别脂肪酸、氧化低密度脂蛋白、晚期氧化终产物及凋亡细胞等配体,参与脂质摄取、免疫识别与调节、炎症反应、血管生成、细胞黏附及凋亡等多个病理生理过程。CD36的表达受转录和翻译后修饰等多个环节的调控。在巨噬细胞、单核细胞、树突状细胞、T细胞和B细胞等细胞类型中,CD36作为免疫信号传导与脂肪酸代谢转运的双重受体,发挥关键作用。由于代谢性疾病与免疫失调之间联系紧密,融合细胞代谢与免疫学的免疫代谢研究逐渐兴起。CD36因其在不同免疫细胞和非免疫细胞中呈现的多重功能,成为连接免疫与代谢的纽带,对糖尿病肾病、代谢功能障碍相关脂肪性肝病、动脉粥样硬化和恶性肿瘤等多种疾病的免疫代谢状态产生显著影响。
关键词 CD36 免疫细胞 免疫代谢
CHEN Jinglei, BAO Hao, LIU Zhihong
National Clinical Research Center for Kidney Diseases, Jinling Hospital, Affiliated Hospital of Medical School, Nanjing University, Nanjing 210016,China
ABSTRACT CD36 is a multifunctional receptor that recognizes ligands such as fatty acids, oxidized low-density lipoprotein, advanced oxidation end products, and apoptotic cells. It plays a role in lipid uptake, immune recognition and regulation, inflammatory responses, angiogenesis, cell adhesion, and apoptosis. The expression and intracellular localization of CD36 are regulated by various ligands through transcriptional and post-translational modifications. CD36 functions as a critical receptor for immune signaling, fatty acid metabolism, and transport in diverse cells, including macrophages, monocytes, dendritic cells, T cells, and B cells. The close association between metabolic diseases and immune dysregulation has led to the development of immunometabolism, a research field that integrates cellular metabolism and immunology. Due to its multifaceted roles in different immune and non-immune cells, CD36 serves as a bridge connecting immunity and metabolism, significantly influencing the immunometabolic status in various diseases, such as diabetic nephropathy, metabolic dysfunction-associated steatotic liver disease, atherosclerosis, and cancer.
Key words CD36 immune cells immunometabolism
近年来,免疫代谢及其在肿瘤和自身免疫病中的作用机制逐渐成为新兴的研究领域之一。CD36是广泛表达于髓系固有免疫细胞等多种细胞表面的跨膜糖蛋白,既是病原相关分子模式(PAMP)和损伤相关分子模式(DAMP)的信号受体,又是长链游离脂肪酸转运蛋白,介导脂质摄取、免疫识别和调节、炎症反应、血管生成、分子黏附和细胞凋亡等过程[1]。由于具有信号传导受体和脂肪酸吸收分子的双重作用,CD36是调节免疫细胞代谢和功能的关键分子。本文综述了CD36在多种免疫细胞中的作用,以及多种疾病中CD36免疫和代谢功能,并对该领域存在的问题进行了展望。
CD36亦称脂肪酸转位酶(FAT)、血小板糖蛋白Ⅳ(GP4)或清道夫受体B2(SR-B2),是表达于血小板、单核细胞、巨噬细胞、脂肪细胞等多种细胞表面的多功能受体,在脂质代谢与天然免疫中扮演关键角色,参与脂质摄取、免疫识别与调节、炎症反应等过程。
人CD36基因位于染色体7q11.2,全长约46 kb。CD36基因突变不仅可导致GP4缺乏症,还与血浆脂肪酸和三酰甘油异常有关,是胰岛素抵抗、代谢性疾病的危险因素[2]。CD36转录受过氧化物酶体增殖物激活受体(PPAR)α和γ调控。研究表明脂肪酸、高葡萄糖、胰岛素、巨噬细胞集落刺激因子、肿瘤坏死因子α(TNF-α)和白细胞介素(IL)-4等均可促进CD36在单核细胞和巨噬细胞等细胞中上调[3]。
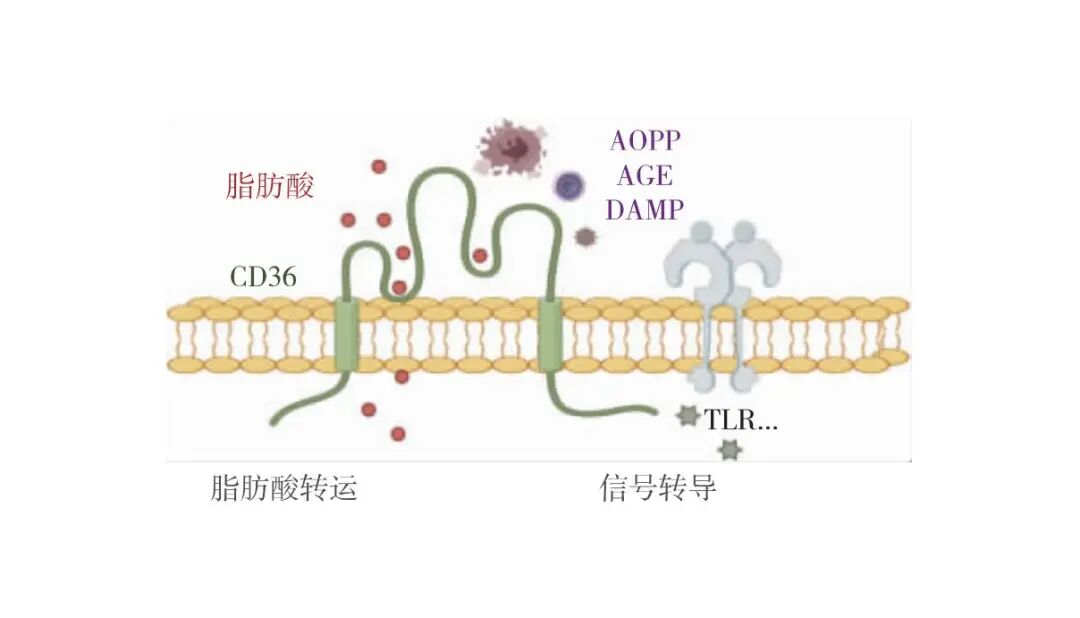
人CD36蛋白含有472个氨基酸,分子量53 kD,具有2个跨膜区和1个含配体结合位点的大胞外区,2个末端各有一条短胞质尾巴。CD36的2个短胞内结构域(N端7个氨基酸,C端13个氨基酸)缺乏激酶/磷酸酶活性及支架域,需通过招募Toll样受体(TLR)2/4/6、Na+/K+ATP酶、整合蛋白及跨膜蛋白等信号复合物传递信号[4]。CD36的胞外区形成2个大型疏水空腔,可介导脂肪酸、磷脂和胆固醇等疏水分子吸收。CD36胞外区还有一个带正电荷的赖氨酸簇区,可结合带负电荷的配体,如晚期氧化蛋白产物(AOPP)、晚期糖基化终末产物(AGE)及凋亡细胞等[5]。这些蛋白作为氧化应激和脂质/蛋白质变性的标志,与CD36的结合能触发炎症等病理生理反应,在多种代谢性疾病、癌症定植转移以及各类肾脏损伤中扮演关键角色[6]。CD36也作为模式识别受体(PRR),识别和吞噬病原体以及 PAMP和 DAMP,促进炎症因子释放(图1)。CD36蛋白还可发生糖基化、泛素化和棕榈酰化等蛋白质翻译后修饰,调节其稳定性、蛋白质折叠和运输。细胞外位点的磷酸化也影响其对脂肪酸等的摄取[7]。
图1 CD36作为信号转导和脂肪酸转运蛋白的多功能受体
AOPP:晚期氧化蛋白产物;AGE:晚期糖基化终末产物;DAMP:损伤相关分子模式;TLR:Toll样受体;本图由Figdraw绘制
可溶性CD36(sCD36),由暴露的跨膜CD36的胞外区域构成,由血浆蛋白酶切割形成或由特定的循环微粒亚群脱落形成[8]。现有研究发现系统性红斑狼疮(SLE)等疾病中sCD36水平升高,sCD36致病机制值得进一步研究。
巨噬细胞 作为在一些细菌、酵母和寄生虫上表达的PAMP的受体,巨噬细胞CD36参与对感染的先天免疫反应。当巨噬细胞CD36表达上调时,长链脂肪酸在线粒体中积聚并促进活性氧(ROS)的产生,线粒体ROS随后激活核因子κB(NF-κB),促进巨噬细胞炎症激活[9]。另一项研究称肿瘤相关巨噬细胞也会增量表达CD36以增加长链脂肪酸吸收,导致细胞内脂质积聚和更高的线粒体脂肪酸β氧化(FAO)。FAO的升高促进了线粒体氧化磷酸化和ROS的产生,激活下游基因转录并促进肿瘤发生[10]。在小鼠中饮食诱导的肥胖显著增强脂肪组织巨噬细胞的炎症激活并促进氧化应激,而CD36缺失的小鼠脂肪组织巨噬细胞和脂肪细胞的炎症反应和胰岛素抵抗降低[11]。
循环单核细胞 CD 36在循环单核细胞中的作用尚不清楚。Lee等[12]证明高表达CD36的单核细胞的抗原呈递可能有助于新生儿和成人中具有调节功能的Foxp3+ T-bet+ T细胞的外周发育。而且在动脉粥样硬化早期,CD36通过调节巡逻活性对非经典单核细胞的功能产生重要影响,CD36缺失导致氧化型低密度脂蛋白(ox-LDL)的摄取降低,最终降低细胞巡逻的速度和距离。CD36作为清道夫受体,还参与单核细胞抗磷脂抗体的活化,与抗磷脂综合征患者的血栓形成相关[13]。此外,Reiss等[14]发现SLE患者血浆会以剂量依赖的方式显著刺激人单核细胞(THP-1)和巨噬细胞中CD36表达升高。
树突状细胞 CD36在树突状细胞(DC)亚群上也高度表达,包括“不成熟”DC和CD8+DC。研究表明,小鼠DC中关键自噬基因的缺失与CD36表达增加、内化肿瘤细胞的能力提升、细胞内脂质积聚增加和CD4+T细胞激活减少有关[15]。在另一项研究中,高谷氨酸钠血症小鼠模型中分离的DC细胞内游离脂肪酸增加,活性氧增加,细胞程序性死亡配体1(PD-L1)和转化生长因子β(TGF-β)的表达增加[16]。还有研究发现,AGE可能通过CD36影响DC和单核细胞对食物过敏原的识别、摄取和抗原加工。这些研究表明CD36介导的脂质积聚可以改变DC代谢,并促进T细胞和自然杀伤细胞免疫抑制。
T细胞 在癌症免疫治疗的背景下,CD36在T细胞代谢和功能中的作用引起了广泛关注。CD36可介导细胞表面抗原的转移,促进胸腺Treg细胞受体库发育和同种异体耐受[17]。Wang等[18]发现,在人类癌症和小鼠肿瘤模型中,肿瘤内Treg细胞中CD36选择性上调并促进其抑制活性;用单克隆抗体靶向CD36还可减弱Treg介导的免疫抑制并增强抗肿瘤免疫。机制上,肿瘤内Treg细胞CD36表达增高导致脂肪酸摄入增加,从而刺激PPAR-β信号传递,通过电子传递链持续产生烟胺腺苷二核苷,将L-乳酸代谢为丙酮酸,以使Treg细胞适应高乳酸肿瘤微环境(TME),获得更好的存活和免疫抑制功能[19]。
B细胞 研究发现CD36可介导B细胞自噬,并与自噬体膜蛋白共定位,CD36缺陷会显著减少B细胞中浆细胞的形成和激活[20]。而脾脏成纤维细胞网状细胞衍生的乙酰胆碱,通过CD36促进脂质内流和自身反应性B细胞的产生;CD36介导的脂质内流可促进SLE时B细胞线粒体氧化磷酸化和浆细胞分化[21]。B细胞特异性敲除 CD36的小鼠会在体内表现出生发中心B细胞和抗DNA抗体的减少。通过质谱、免疫荧光和免疫共沉淀等技术证实,CD36与抑制性Fc受体 FcγRⅡb 之间的相互作用,小鼠中 FcγRⅡb的缺失会导致边缘区 B 细胞、生发中心B细胞和浆细胞中CD36的表达降低,进一步证实了CD36在B细胞代谢和体液反应中的重要作用[22]。这些研究表明,CD36参与B细胞关键脂肪酸代谢、生物过程,进而影响B细胞激活和生存/死亡状态。
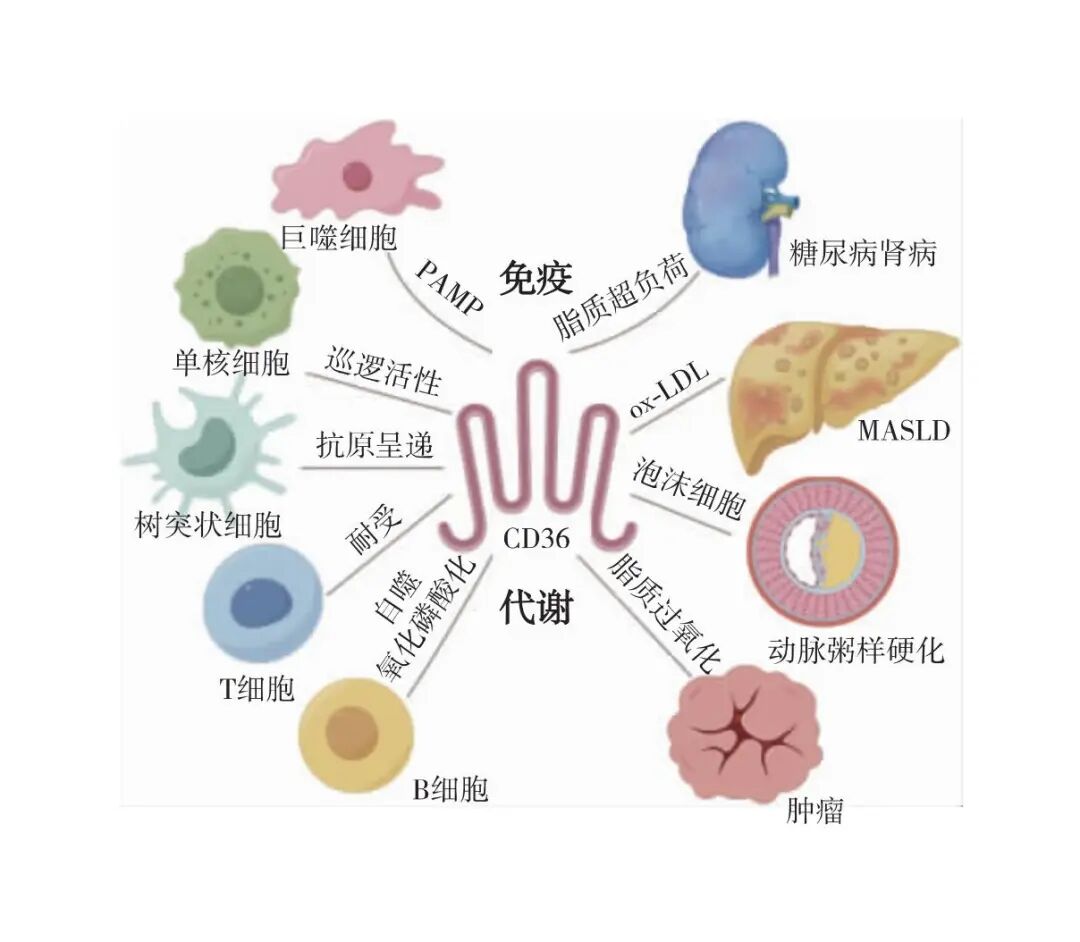
“免疫代谢”由Mathis和Shoelson最先提出,他们观察到代谢性疾病通常与免疫系统失调有关[23]。免疫代谢是集细胞代谢和免疫学于一体的新兴研究领域,随着近来研究的突破性进展,逐渐认识到细胞代谢不仅仅是在免疫细胞激活期间DNA、RNA、蛋白质和脂类等大分子的重新合成提供能量和构建基础,事实上许多代谢中间体本身就是导致促炎或抗炎反应的关键信号分子。类似于免疫细胞活化的动态性,细胞代谢途径是高度灵活的,因为免疫细胞面对不同的细胞外环境会发生代谢重编程,这通常先于甚至驱动免疫细胞的分化、活化和死亡。免疫代谢领域的一个关键问题是细胞代谢途径和免疫细胞分化/激活是如何整合的,以及这种整合在病理条件下是如何失调的。CD36既是免疫细胞上PAMP/DAMP等的受体,可以激活下游的不同信号通路,触发炎症等病理生理反应;又是细胞吸收长链脂肪酸、ox-LDL和磷脂等的转运体(图2)。
图2 CD36在免疫代谢中的作用
PAMP:病原相关分子模式;ox-LDL:氧化型低密度脂蛋白;MASLD:代谢功能障碍相关脂肪性肝病;本图由Figdraw绘制
高血脂和糖尿病肾病 内皮细胞CD36已被证明可介导脂肪酸摄入心脏、骨骼肌和棕色脂肪组织,从而影响葡萄糖利用和胰岛素敏感性。内皮细胞CD36还可调节膜神经酰胺形成、外泌体脂肪酸转移和循环脂肪酸水平。在肾脏中,糖尿病肾病患者发生心血管事件的风险显著升高与脂代谢紊乱密切相关[24]。由于线粒体FAO是肾脏ATP的主要来源,在近端肾小管上皮细胞中CD36相关的脂肪酸线粒体转移为FAO的限速步骤,相关FAO缺陷引起细胞能量重编程,糖酵解比例增加,细胞内脂质沉积增加,最终导致肾间质纤维化。糖尿病肾病肾小管上皮细胞CD36表达增强,通过使腺苷酸活化蛋白激酶(AMPK)信号通路失活,导致FAO减少和糖酵解增加,并促进线粒体ROS的产生,从而激活NLR家族Pyrin域蛋白3(NLRP3)炎症体信号通路,导致疾病进展[25]。还有研究发现酰基蛋白硫酯酶1(APT1)可在足细胞中使CD36去棕榈酰化,糖尿病诱导的APT1缺乏通过促进质膜上棕榈酰化的CD36富集,最终导致脂质超负荷和足细胞损伤[26]。
代谢功能障碍相关脂肪性肝病(MASLD) MASLD是一种代谢性肝病,其特征是肝细胞内大量脂肪堆积。MASLD 的早期阶段是单纯性脂肪肝(肝脂肪变性),更严重的阶段就是代谢功能障碍相关脂肪性肝炎(MASH),最终进展为肝纤维化、肝硬化和肝癌。CD36会增加肝脏中脂肪酸的摄取,过量的脂肪酸内流导致肝细胞氧化应激和过氧化,从而导致脂质紊乱,进而诱导肝细胞的细胞毒性、线粒体自噬和细胞凋亡,促进组织损伤和炎症,从而驱动肝脂肪变性发作和最终进展为MASH[27]。Bieghs团队[28]证实肝脏巨噬细胞CD36吞噬修饰脂质(例如ox-LDL),导致胆固醇在溶酶体中积累并引起肝脏炎症,从而增加MASH的风险。CD36的信号传导还通过介导线粒体代谢重编程,从氧化磷酸化转变为超氧化物的产生,响应其配体ox-LDL,驱动巨噬细胞NF-κB活化和炎症因子生成[9]。
动脉粥样硬化 动脉粥样硬化是动脉壁的一种慢性进行性炎症,表现为泡沫细胞积累、巨噬细胞滞留在斑块中和血栓形成。泡沫细胞生成的主要原因是ox-LDL的过度流入,以及胆固醇酯在内膜巨噬细胞中的积累。作为巨噬细胞中重要的清道夫受体,CD36在细胞ox-LDL积累和泡沫细胞形成中起关键作用。由于巨噬细胞具有高亲和力的CD36相关oxLDL,因此通过CD36与ox-LDL结合并内化ox-LDL[29]。随着CD36-oxLDL组装体的内化,ox-LDL进入细胞并积累,将巨噬细胞转移到泡沫细胞中。除了泡沫细胞的形成外,CD36还参与巨噬细胞的炎症反应。例如,靶向CD36会抑制NLRP3炎性小体,导致动脉粥样硬化小鼠血清IL-1β降低[30]。CD36相关ox-LDL摄取激活巨噬细胞氧化应激标志物和炎症标志物的特异性表达谱。即使是相对低浓度的ox-LDL也可通过表观遗传组蛋白修饰诱导持久的致动脉粥样硬化的巨噬细胞表型[31]。
肿瘤免疫 肿瘤组织形成需要特定的TME。除了肿瘤细胞,TME中还有免疫细胞、基质细胞和其他细胞。TME的一个表征是脂代谢紊乱和失调,增高的脂类吸收(包括脂肪酸、胆固醇)和细胞内脂质富集,不但抑制抗肿瘤的免疫应答,同时增强促肿瘤的免疫应答。研究发现肿瘤环境中的胆固醇可诱导CD8+T细胞CD36表达的升高,进而摄取过多的脂肪酸,引发脂质过氧化和铁死亡,导致其杀伤性功能的丧失,促进肿瘤的生长;靶向CD36和铁死亡可能是提高CD8+T细胞免疫抗肿瘤疗法的有效策略[32]。肿瘤浸润CD8+T细胞上调CD36的表达水平,并利用其吸收Ox-LDL,诱导脂质过氧化和p38激酶下游活化从而抑制CD8+T细胞的效应功能[33]。CD36作为外源性脂质的转运蛋白,也可促进急性髓性白血病(AML)免疫逃避。除了在脂质氧化中的既定作用外,AML 细胞上的 CD36可感应Ox-LDL以激活TLR4-LYN-MYD88-NF-κB通路,并且通过 CD36的外源棕榈酸酯转移促进ZDHHC6介导的MYD88棕榈酰化,进一步增强这种先天免疫通路[34]。这些都说明CD36通过对脂质代谢的调节影响免疫细胞的功能和活化状态以及肿瘤的发生和发展。
此外,还有研究证明CD36在多种疾病中都通过改变免疫代谢充当其发病中不可缺失的一环,如感染、肺动脉高压、视网膜炎症等。
CD36是一种多功能受体,能够识别多种配体,包括脂肪酸、ox-LDL、终末生物氧化产物(如AOPP、AGE)及凋亡细胞等。CD36是联系炎症与细胞代谢的关键桥梁。通过在各种免疫和非免疫细胞中充当DAMP/PAMP受体,CD36介导了细胞内信号传导、ROS产生和转录因子激活等促进炎症;同时CD36还是长链脂肪酸等的转运体。由于CD36在免疫信号传导和脂肪酸代谢中扮演着多重角色,这使得CD36在多种疾病,如糖尿病肾病、MASLD、动脉粥样硬化和恶性肿瘤中,对免疫代谢状态产生重要的影响。CD36是一个可能的治疗干预靶点,但其生物学功能复杂,仍存在许多问题,包括信号传导和长链脂肪酸运输的双重功能如何协调和整合?在表达CD36的细胞中,这种双重功能在代谢调节和细胞分化、活化及存活中的作用有多大?这些都值得我们进一步探究。
参考文献
【引用本文】陈晶蕾、鲍浩、 刘志红. CD36 在免疫代谢中的作用[J]. 肾脏病与透析肾移植杂志, 2025, 34(6): 570-574.
CHEN Jinglei, BAO Hao, LIU Zhihong. The role of CD36 in immunometabolism[J]. Chinese Journal of Nephrology, Dialysis & Transplantation, 2025, 34(6): 570-574.
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017