



200
评论
查看更多
密码过期或已经不安全,请修改密码
修改密码
壹生身份认证协议书
同意
拒绝
同意
拒绝
同意
不同意并跳过





上期回顾↓↓
作者:安徽医科大学第一附属医院消化内科 徐桂芳 王守宇
IBD患者菌群多样性显著下降,表现为专性厌氧菌(如Faecalibacterium prausnitzii)减少,兼性厌氧菌(如肠杆菌科)扩增。黏附浸润型大肠杆菌(AIEC)是CD最具代表性的致病共生菌,约30%的CD患者肠道可检出。AIEC的致病三步骤为:①通过FimH蛋白结合肠上皮CEACAM6;②携带PTPN2 rs1893217风险等位基因者,JAK-STAT通路过度激活,CEACAM6表达进一步升高,AIEC侵袭增强,JAK抑制剂托法替尼可逆转该效应;③AIEC进入巨噬细胞后通过“两阶段适应策略”形成胞内细菌群落(IBCs)-先进入持留状态,随后恢复复制并嵌入生物膜样基质,耐受溶酶体应激与抗生素,持续驱动炎症[2-5]。
(1)短链脂肪酸:丁酸盐不仅为肠上皮供能,更通过抑制组蛋白去乙酰化酶3(HDAC3)诱导组蛋白H3K9高乙酰化,显著降低IBD肠黏膜TNF-α和IL-6分泌,并抑制巨噬细胞CD40/CD80表达[6]。同时,丁酸盐可选择性地将AIEC吞噬率提高62%,而对共生菌吞噬下降,体现精准清除致病菌的能力。在肠上皮细胞中,丁酸盐主要恢复炎症条件下下调的MCT1(自身转运体)及IL-18表达,促进屏障修复,但对上皮细胞本身的抗炎作用有限。
(2)次级胆汁酸:肠道菌群可将初级胆汁酸转化为多种具有免疫调节活性的次级胆汁酸。其中,3-氧代石胆酸(3-oxoLCA)能够直接结合并拮抗转录因子RORγt的活性,从而抑制致病性Th17细胞的分化。石胆酸衍生物isoalloLCA则通过诱导活性氧产生,上调Foxp3的表达,促进Treg细胞的分化。正常情况下,这两类次级胆汁酸共同维持Th17/Treg的免疫平衡[7]。然而,IBD患者肠道中产生这些胆汁酸的细菌丰度显著降低,导致3-oxoLCA和isoalloLCA生成减少,使得Th17/Treg天平向促炎方向倾斜,从而加剧肠道炎症。
肠道黏液层O-聚糖结构为细菌提供锚点。IBD中糖基化酶异常导致黏液层变薄紊乱,微生物抗原直接接触上皮下树突状细胞,打破隔绝,促进免疫识别,加剧炎症反应。
粪菌移植(FMT)、益生菌、噬菌体疗法及饮食干预(如补充可溶性纤维)等用于IBD的治疗方案正在探索中。AIEC靶向噬菌体混合物EcoActive在临床前模型中显示出特异性清除AIEC的潜力。
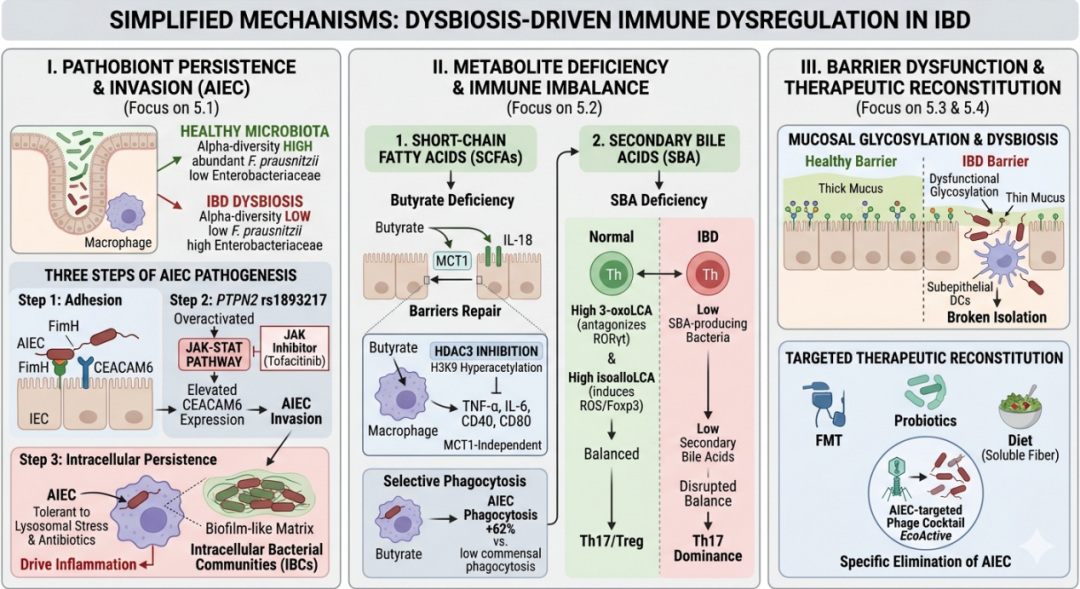
图3. 肠道菌群与免疫互作
随着研究的深入,人们发现,表观遗传调控深刻影响着IBD的进展。研究表明,IBD患者肠上皮CEACAM6启动子区甲基化异常受HIF-1α调控;HDAC3抑制剂(如丁酸盐)通过组蛋白高乙酰化抑制炎症;长链非编码RNA GAS5通过增加小穹隆RNA水平破坏上皮屏障,促进IBD进展。
肠道神经系统与免疫系统之间存在精密的双向通讯,共同维持黏膜稳态。其中,降钙素基因相关肽(CGRP)家族成员发挥着关键的免疫调节作用。最新研究发现,肠道内源性感觉神经元产生的肾上腺髓质素2(ADM2)可通过作用于2型固有淋巴细胞(ILC2)表面的特异性受体(CALCRL-RAMP3复合物),显著促进ILC2产生组织保护性因子双调蛋白(AREG),从而限制肠道炎症并促进黏膜修复[8]。在IBD患者中,ADM2及其受体亚基CALCRL和RAMP3的表达显著改变,且与ILC2中AREG表达降低相关。这些发现揭示了神经肽ADM2通过调控ILC2功能发挥组织保护作用的新机制,为IBD的神经免疫治疗提供了潜在新靶点。
IL-17REL是IL-17家族新成员,可作为诱饵受体负向调控IL-17信号,其功能异常可能与IBD相关。其他新型免疫检查点如TIGIT、VISTA在IBD中的作用尚在探索中。
外泌体在IBD诊疗中展现出双重潜力。作为非侵入性生物标志物,唾液外泌体来源的8-miRNA特征谱可高效区分IBD患者与健康对照(AUC达0.925)[9]。在治疗层面,间充质干细胞(MSC)来源外泌体通过递送miR-34a-5p或TSG-6蛋白,分别抑制炎症并促进黏膜愈合[10]。此外,植物来源外泌体(如黄精PSELNs)携带miR166u靶向CD11c,通过TLR4/AKT通路减轻结肠炎[11]。
细胞焦亡是IBD急性炎症的核心驱动机制:GSDMD被炎症小体(如NLRP3、AIM2)切割后在细胞膜上成孔,释放IL-1β和IL-18,加剧屏障损伤[12]。具核梭杆菌外泌体携带致病DNA激活AIM2炎症小体,诱导肠上皮细胞焦亡[13]。天然化合物PEITC通过阻断TXNIP-NLRP3相互作用抑制焦亡,而中草药成分imperatorin则靶向AIM2通路减轻肠纤维化[14]。靶向GSDMD的小分子抑制剂及工程化外泌体(如TSG-6修饰)正进入临床前评估,为控制IBD急性炎症提供新策略。
(1)抗TNF制剂:英夫利西单抗、阿达木单抗仍是IBD治疗的基石,但约30%~40%患者原发无应答或继发失应答。研究显示,CD患者存在FOXP3⁺促炎性Treg亚群[15],该亚群对TNF-α信号异常响应且抑制功能受损,而组蛋白去乙酰化酶(HDAC)抑制剂伏立诺他(vorinostat)可逆转其异常基因表达谱,恢复Treg的抑制功能,为抗TNF治疗失败者提供了新的联合治疗策略。
(2)整合素拮抗剂:维得利珠单抗(抗α4β7)选择性抑制肠道淋巴细胞归巢,已在临床广泛应用。
(3)JAK抑制剂:托法替尼、乌帕替尼通过抑制JAK-STAT通路阻断多种细胞因子信号。托法替尼还可通过逆转PTPN2风险等位基因介导的CEACAM6过表达,限制AIEC侵袭。
(4)其他小分子靶向药物:最新研究揭示,传统的钙神经蛋白抑制剂他克莫司可直接修复肠上皮屏障[16]。此外,伏立诺他作为HDAC抑制剂,除调节Treg功能外,还可抑制DSS诱导的结肠炎,其口服制剂正在CD临床试验中评估[17]。
(5)选择性B细胞靶向治疗:利妥昔单抗在IBD中疗效不明确,但针对浆细胞或BAFF/APRIL通路的策略值得探索。
间充质干细胞(MSC)及其外泌体通过递送抗炎分子(如TSG-6、miR-34a-5p)抑制NLRP3/GSDMD轴,减轻结肠炎[18]。植物外泌体样纳米颗粒(如黄精来源PSELNs)也显示出治疗潜力。
研究表明,约40%患者对现有治疗应答不佳,亟需基于分子分型的精准治疗。唾液外泌体miRNA、血清抗AIEC抗体等无创标志物正在验证中,而靶向菌群-免疫互作的多维度干预(饮食、益生菌、噬菌体、FMT)有望填补单靶点药物的不足。
患者来源肠上皮类器官(PDOs)可在体外长期扩增并保留原始组织的转录组特征,与体内肠上皮约90%相似。CD患者PDOs仍保留疾病表型特异性的基因表达谱和分泌组特征,可用于筛选逆转表型的候选化合物。优化的成体干细胞来源人小肠类器官可同时分化为所有肠上皮细胞类型,并揭示IL-22通过PI3K/AKT/mTOR轴诱导潘氏细胞分化的新机制[19-20]。人诱导多能干细胞来源结肠类器官(hiPSC-COs)包含上皮和间质细胞,经TNF-α、IFN-γ和IL-1β处理可重现UC的炎症反应和屏障损伤,用于评估托法替尼等药物疗效[21]。
精密切割肠片(PCIS)作为一种新兴的离体模型,保留了肠道完整的三维结构和驻留免疫细胞,能够模拟IBD组织微环境,可用于抗炎药物的快速筛选和机制验证[22]。这些模型创新为IBD基础研究向精准治疗转化提供了重要平台。
综上,IBD免疫学基础研究已从“单一细胞/通路”转向“细胞-微生物互作/免疫-代谢-表观网络调控”,未来需整合多组学及活体成像技术,在单细胞精度上解析免疫网络的时空动态,加速基础研究向精准干预的临床转化。

安徽医科大学第一附属医院消化科主任
主任医师,教授,博士生导师
安徽省医师协会消化内镜学分会主任委员
中华医学会消化内镜学分会ESD协作组副组长
中华医学会消化内镜学分会显微内镜协作组委员
中华医学会消化病学分会激素与黏膜学组委员
安徽省消化内镜微创诊疗专科联盟理事长
安徽省合肥市蜀山区妇联副主席
《中华消化内镜杂志》通讯编委
《中华消化杂志》青年编委
安徽省江淮英才项目创新领军人才
安徽医科大学第一附属医院教育部肿瘤免疫医药基础研究创新中心,教授、博士生导师
安徽省领军人才特聘教授
安徽省抗癌协会整合肿瘤研究专委会 副主任委员
安徽省抗癌协会免疫与基因诊断专委会 副秘书长
江苏省杰出青年基金获得者
美国 M.D. Anderson 癌症中心访问学者
[1] Horn V, Sonnenberg GF. Group 3 innate lymphoid cells in intestinal health and disease. Nat Rev Gastroenterol Hepatol. 2024;21:349-364.
[2] Chatterjee P, Canale V, et al. The PTPN2 rs1893217 IBD risk allele increases susceptibility to AIEC invasion by a JAK-STAT-CEACAM6 axis. Gut Microbes. 2025;17(1):2526136.
[3] Stankey CT, Bourges C, et al. A disease-associated gene desert directs macrophage inflammation through ETS2. Nature. 2024;630(8016):447-456.
[4] Bruder E, Nedjar H, et al. Persistence mechanisms of Crohn‘s disease-associated adherent invasive Escherichia coli within macrophages. Gut Microbes. 2025;17(1):2587402.
[5] Barnich N, Arthur JC, et al. Adherent-invasive Escherichia coli in Crohn’s disease: the 25th anniversary. Gut. 2025;75(2):411-424.
[6] Parada-Venegas D, De la Fuente M, et al. Butyrate suppresses mucosal inflammation in inflammatory bowel disease primarily through HDAC3 inhibition in monocytes and macrophages. FEBS J. 2025;292(22):6134-6157.
[7] Paik D, Yao L, et al. Human gut bacteria produce ΤΗ17-modulating bile acid metabolites. Nature. 2022;603:907-912.
[8] Uddin J, Yano H, et al. CGRP-related neuropeptide adrenomedullin 2 promotes tissue-protective ILC2 responses and limits intestinal inflammation. Nat Immunol. 2025;26(9):1516-1526.
[9] Yang C, Chen J, et al. Identification of Salivary Exosome-Derived miRNAs as Potential Biomarkers for Non-Invasive Diagnosis and Proactive Monitoring of Inflammatory Bowel Disease. Int J Mol Sci. 2025;26(16):7750.
[10] Wu B, Su S, et al. Bone marrow mesenchymal stem cell-derived exosomes alleviate DSS-induced inflammatory bowel disease in mice through inhibiting intestinal epithelial cell pyroptosis via delivery of TSG-6. Front Immunol. 2025;16:1601591.
[11] Wei C, Chen Y, et al. miR166u-enriched Polygonatum sibiricum exosome-like nanoparticles alleviate colitis by improving intestinal barrier through the TLR4/AKT pathway. Int J Biol Macromol. 2025;318(Pt 1):144802.
[12] Wang J, Zhang C, et al. Direct inhibition of the TXNIP-NLRP3-GSDMD pathway reduces pyroptosis in colonocytes and alleviates ulcerative colitis in mice by the small compound PEITC. Acta Pharmacol Sin. 2025;46(9):2436-2449.
[13] Li S, Zhu F, et al. Imperatorin Alleviates Intestinal Fibrosis by Suppressing AIM2-mediated GSDMD Pyroptosis in Macrophages. Cell Mol Gastroenterol Hepatol. 2025;101625.
[14] Zhao Y, Wu R, et al. Fusobacterium nucleatum-derived extracellular vesicles carrying virulence factor DNA trigger AIM2 inflammasome activation to facilitate UC progression. Cell Mol Biol Lett. 2025;30:138.
[15] Kosinsky RL, Gonzalez M, et al. The FOXP3⁺ pro-inflammatory T cells in Crohn‘s disease: a potential therapeutic target. Gastroenterology. 2024;166(4):634-644.e14.
[16] Zuo L, Kuo WT, et al. Tacrolimus-binding protein FKBP8 directs myosin light chain kinase-dependent barrier regulation and is a potential therapeutic target in Crohn’s disease. Gut. 2023;72(5):870-881.
[17] Villablanca EJ, Selin K, Hedin CRH. Mechanisms of mucosal healing: treating inflammatory bowel disease without immunosuppression? Nat Rev Gastroenterol Hepatol. 2022;19:493-507.
[18] Lee Y, Yoo C, et al. Immunoregulatory protein-hybrid extracellular vesicles via self-loadable backbone cyclization for oral inflammatory bowel disease therapy. Bioact Mater. 2026;58:70-88.
[19] Niklinska-Schirtz BJ, Venkateswaran S, et al. Ileal Derived Organoids From Crohn‘s Disease Patients Show Unique Transcriptomic and Secretomic Signatures. Cell Mol Gastroenterol Hepatol. 2021;12(4):1267-1280.
[20] He GW, Lin L, et al. Optimized human intestinal organoid model reveals interleukin-22-dependency of paneth cell formation. Cell Stem Cell. 2022;29(9):1330-1343.
[21] Yokoi F, Deguchi S, et al. Establishment of an ulcerative colitis model using colon organoids derived from human induced pluripotent stem cells. iScience. 2024;27(10):111049.
[22] Beaurivage C, et al. Development of a human primary gut-on-a-chip to model inflammatory processes. Sci Rep. 2020;10:21475.
【安医心法】CD规范化诊断 | IBD诊疗能力系统提升2

本文版权属于中国医学论坛报社,转载须授权
查看更多
 中国医学论坛报
中国医学论坛报 壹生
壹生 今日肿瘤
今日肿瘤 今日循环
今日循环 今日糖尿病
今日糖尿病 今日口腔
今日口腔 全科周刊
全科周刊 脱贫地区农副产品网络销售平台
脱贫地区农副产品网络销售平台
京公网安备 11010202008182号
| 互联网新闻信息服务许可证编号:10120190017